La reproduction, caractéristique fondamentale de tout être vivant, est intrinsèquement liée à la capacité des organismes à augmenter leur nombre de cellules. Chez les plantes, tout comme chez les animaux, ce processus est orchestré par la division cellulaire, un mécanisme d'une complexité fascinante qui a évolué au fil du temps pour garantir une transmission fidèle du matériel génétique et une croissance organisée. Alors que les formes de vie les plus primitives, comme les bactéries, utilisent la scissiparité, un mode de division simple et rapide, les organismes eucaryotes, tels que les plantes supérieures, ont développé des processus plus élaborés, au premier rang desquels figure la mitose. Cette dernière assure une parité génétique parfaite entre les cellules filles et joue un rôle crucial dans le développement et le maintien de la structure végétale.
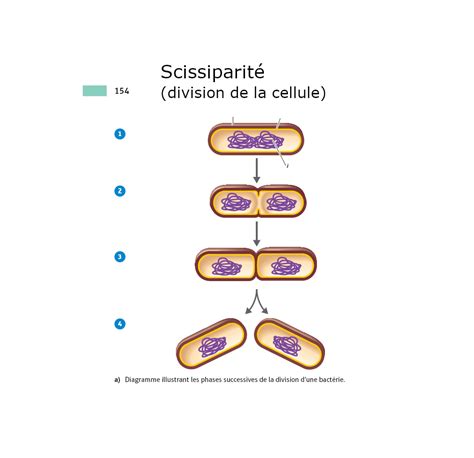
Le Cycle Cellulaire : Une Préparation Active à la Division
L'intervalle de temps entre deux divisions cellulaires successives, appelé cycle cellulaire, n'est pas une simple période de repos. Au contraire, il s'agit d'une phase d'intense préparation, divisée en trois étapes clés : G1, S et G2.
La phase G1 suit immédiatement la mitose. Les cellules filles, de taille réduite, entament une période de croissance active. Durant cette phase, la cellule augmente sa taille, ses organites se multiplient, et la chromatine, le matériel génétique sous forme décondensée, commence à se détendre. C'est également durant la phase G1 que la cellule franchit un point de décision crucial, le point de restriction (R). Ce seuil est déterminant : une fois franchi, la division cellulaire devient irréversible, garantissant que le processus se déroulera jusqu'à son terme, quelles que soient les conditions environnementales ou internes. Le franchissement de R peut être retardé, permettant ainsi à la cellule d'allonger son cycle et de s'adapter.
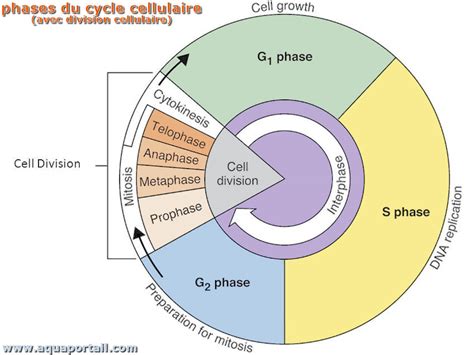
La phase S est marquée par la réplication de l'ADN. La cellule duplique son matériel génétique, assurant que chaque future cellule fille recevra une copie complète et identique du génome. La fin de cette phase est également un repère temporel important.
Enfin, la phase G2 représente la dernière étape de préparation avant la mitose. La réplication de l'ADN est achevée, et la cellule synthétise les molécules nécessaires à la division, tout en condensant sa chromatine pour former des chromosomes individualisés et visibles au microscope.
Le Positionnement de la Paroi : Une Contrainte Spatiale chez les Végétaux
Une caractéristique distinctive des plantes est leur immobilité. Cette sédentarité se manifeste également au niveau cellulaire : les cellules végétales ne migrent pas, restant soudées les unes aux autres par la paroi pecto-cellulosique. Cette rigidité structurelle confère une importance capitale au positionnement de la nouvelle paroi qui se formera lors de la division.
Le cytosquelette joue un rôle prépondérant dans la détermination de cet emplacement. Au début de la prophase mitotique, les microtubules, initialement répartis uniformément sous le plasmalemme, se concentrent pour former une "bande préprophasique" le long d'un diamètre de la cellule. Le noyau se positionne ensuite au centre de cette bande, guidé par des filaments d'actine.
Pendant que le noyau progresse dans les différentes étapes de la mitose, les microtubules se réorganisent pour former le "fuseau achromatique". Ce dernier atteint sa pleine maturité en métaphase, moment où les chromosomes, au maximum de leur condensation, s'alignent sur la "plaque équatoriale".
Après la division du noyau (cytodiérèse), la construction de la nouvelle paroi (cytokinèse) débute. C'est le phragmoplaste, une structure formée de vésicules chargées de pectine issues du Golgi, qui initie cette construction. Ces vésicules fusionnent pour former une lamelle pectique, qui deviendra la paroi séparant les deux cellules filles. Des interruptions dans cette lamelle permettent la formation des plasmodesmes, des canaux de communication intercellulaires.
Les Divisions Cellulaires et la Morphogenèse Végétale
L'orientation de la formation de ces nouvelles parois a des conséquences directes sur la morphologie des organes végétaux.
Divisions périclines : Lorsque les nouvelles parois se forment parallèlement à la surface du tissu, les divisions sont dites périclines. Ce type de division est observé, par exemple, dans le méristème fondamental d'un embryon ou dans le cambium vasculaire, contribuant à l'épaississement des tiges et des racines.
Divisions anticlines : Si les nouvelles parois sont perpendiculaires les unes aux autres, les divisions sont qualifiées d'anticlines. Ce mode de division est crucial pour l'expansion des tissus et la formation de nouvelles structures, comme on peut l'observer lors de l'embryogenèse, au stade globulaire.
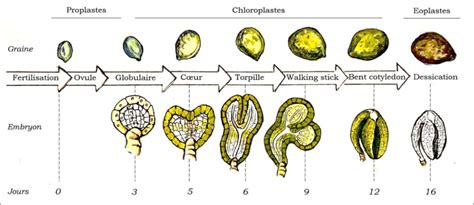
Divisions aléatoires : Lorsque l'orientation des nouvelles parois est aléatoire, la forme de l'organe résultant est également désordonnée.
La Mitose chez les Végétaux : Étapes Clés de la Division Nucléaire
La mitose, ou division du noyau, est un processus continu, mais pour des raisons pédagogiques, elle est traditionnellement décomposée en plusieurs phases distinctes, étudiées ici à partir de coupes de racines de jacinthe colorées pour visualiser l'ADN.
Interphase : Le noyau est intact, délimité par une enveloppe nucléaire. L'ADN est sous forme de chromatine décondensée, et des nucléoles sont visibles. Le cytoplasme est légèrement vacuolisé.
Prophase : La chromatine commence à se condenser, formant des chromosomes individualisés. Les nucléoles et l'enveloppe nucléaire disparaissent progressivement.
Prométaphase : Le fuseau achromatique, composé de microtubules, se forme autour des chromosomes. Des fibres fusoriales s'attachent aux centromères des chromosomes, les préparant à leur déplacement.
Métaphase : Les chromosomes, au maximum de leur condensation, s'alignent sur la plaque équatoriale, au centre du fuseau achromatique.
Anaphase : Les centromères se clivent, séparant les chromatides sœurs. Chaque chromatide, désormais considérée comme un chromosome individuel, est tirée vers l'un des pôles du fuseau par les fibres fusoriales.
Télophase : Les chromosomes atteignent les pôles du fuseau et commencent à se décondenser. Deux nouveaux noyaux se reforment, entourés d'une nouvelle enveloppe nucléaire. Parallèlement, des vésicules s'accumulent à l'équateur pour former le phragmoplaste, précurseur de la nouvelle paroi.
Fin de la Télophase : La plaque cellulaire (phragmoplaste) s'étend, se fusionne avec la paroi existante et sépare définitivement les deux cellules filles. Les chromosomes sont complètement décondensés, et deux noyaux interphasiques sont reconstitués.
Mitosis in an animal cell Under the Microscope
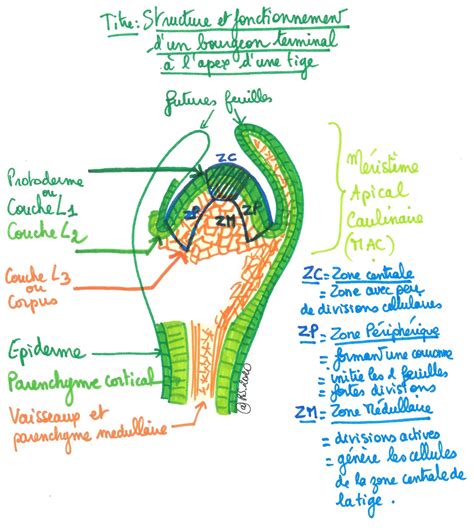
Le Méristème Apical Caulinaire : Un Centre de Croissance Dynamique
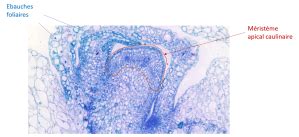
Chez les plantes, la croissance est assurée tout au long de leur vie par des tissus spécialisés appelés méristèmes. Le méristème apical caulinaire (MAC), situé à l'extrémité de la tige, est une structure dynamique responsable de la production de nouvelles cellules qui formeront la tige, les feuilles et les fleurs. Identifié pour la première fois au XVIIIe siècle, son organisation et son fonctionnement ont été précisés au fil des siècles.
Le MAC est organisé en plusieurs couches (L1, L2, L3) et zones fonctionnelles (zone centrale - CZ, zone périphérique - PZ, centre organisateur - OC). Les cellules des couches L1 et L2, qui donneront naissance à l'épiderme et au parenchyme, subissent principalement des divisions anticlines. Dans la couche L3, les divisions peuvent être périclines ou anticlines, contribuant à la formation des tissus vasculaires.
Au cœur de la régulation du MAC se trouve une boucle de rétroaction négative impliquant les cellules souches et le centre organisateur (OC). Le facteur de transcription WUSCHEL (WUS), exprimé dans l'OC, migre vers la zone centrale (CZ) où il maintient les cellules souches et stimule l'expression du peptide de signalisation CLAVATA3 (CLV3). CLV3, à son tour, inhibe l'expression de WUS, régulant ainsi la taille de la population de cellules souches et assurant une croissance équilibrée. Des mutations dans les gènes CLV1, CLV2 ou CLV3 entraînent une hypertrophie du MAC, démontrant l'importance de cette boucle régulatrice.
La signalisation des cytokinines, hormones végétales produites par les racines, joue également un rôle crucial dans la régulation de WUS et, par conséquent, dans la coordination de la croissance entre l'appareil aérien et racinaire. L'équilibre entre les espèces réactives de l'oxygène, comme les ions superoxydes (O2.-) et le peroxyde d'hydrogène (H2O2), agit comme un interrupteur moléculaire pour maintenir l'état de cellule souche ou favoriser la différenciation, influençant ainsi l'expression de WUS.
Régulation du Cycle Cellulaire : Un Mécanisme Sophistiqué
Le cycle cellulaire est finement régulé par un ensemble complexe de protéines, notamment les kinases dépendantes des cyclines (Cdk) et leurs partenaires, les cyclines. Ces complexes Cycline/Cdk agissent comme des interrupteurs moléculaires, déclenchant les événements propres à chaque phase du cycle (G1, S, G2, M).
Régulation de l'activité des Cdk : L'activité des Cdk est strictement contrôlée. Elles sont activées par leur association avec des cyclines spécifiques et par des modifications post-traductionnelles, telles que la phosphorylation et la déphosphorylation de résidus d'acides aminés clés. Des protéines inhibitrices, comme p21, peuvent également bloquer l'activité des complexes Cycline/Cdk.
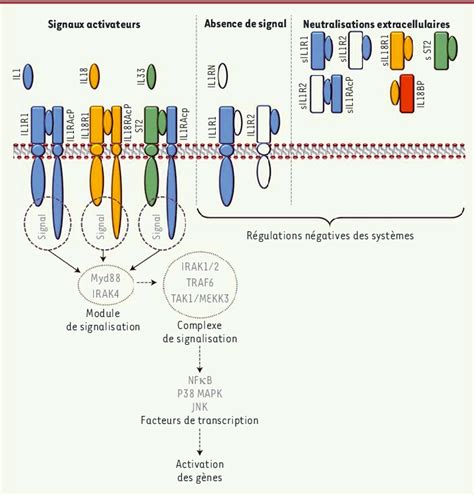
Points de contrôle du cycle : Au-delà de la régulation séquentielle, la cellule dispose de systèmes de surveillance sophistiqués, appelés "points de contrôle" (checkpoints), qui garantissent l'intégrité du génome et la fidélité de la réplication.
- Le DNA Damage Checkpoint (DDCP) détecte les lésions de l'ADN et peut arrêter le cycle en G1, S ou G2 pour permettre les réparations.
- Le Réplication Checkpoint (RCP) s'assure que la réplication de l'ADN est complète avant l'entrée en mitose.
- Le Mitotic Checkpoint (MCP) veille à ce que tous les chromosomes soient correctement attachés au fuseau mitotique avant le déclenchement de l'anaphase, prévenant ainsi des erreurs de ségrégation chromosomique.
Ces mécanismes de surveillance sont essentiels pour prévenir l'accumulation de mutations et sont souvent dérégulés dans les cellules cancéreuses, soulignant l'importance de leur étude pour la compréhension des maladies. La découverte du Maturation Promoting Factor (MPF), un complexe Cycline B/Cdk1, a été une étape majeure dans la compréhension de la régulation de l'entrée en mitose.
La division cellulaire, et plus spécifiquement la mitose chez les plantes, est donc un processus remarquablement orchestré, impliquant une séquence d'événements moléculaires et cellulaires précis. L'organisation spatiale de la formation de la paroi, la régulation rigoureuse du cycle cellulaire et les mécanismes de contrôle sophistiqués garantissent la fidélité de la transmission de l'information génétique et le développement harmonieux de l'organisme végétal.
tags: #division #cellulaire #anticline